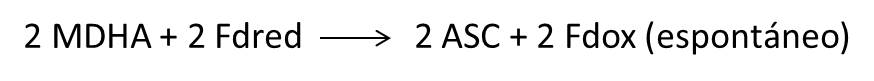La Fotorrespiración: Un mecanismo de protección para la fotosíntesis en condiciones de estrés ambiental
José A. Hernández Cortés (Grupo de Biotecnología de Frutales, CEBAS-CSIC, Murcia)
La RUBISCO (Ribulosa 1,5-bisfosfato Carboxilasa-Oxigenasa) tiene la capacidad de catalizar tanto la carboxilación (ciclo de Calvin-Benson) como la oxigenación de la Ribulosa 1,5-bifosfato (Miziorko y Lorimer 1983). Mientras que la carboxilación de la Ribulosa 1,5-bifosfato da lugar a dos moléculas de 3-fosfoglicerato, la oxigenación produce una molécula de 3-fosfoglicerato y otra de 2-fosfoglicerato. Este proceso de oxigenación de la Ribulosa 1,5 bifosfato se continúa con una serie de reacciones enzimáticas que tienen lugar en tres compartimentos celulares: cloroplastos, peroxisomas y mitocondrias. Por eso no es raro ver a estos tres orgánulos muy próximos el uno del otro en una célula vegetal en hojas (Fig 1). A este conjunto de reacciones se le conoce con el nombre de fotorrrespiración (o ciclo fotosintético C2 de oxidación del carbono) y ocurre fundamentalmente en las plantas denominadas C3 (Ogren 1984).

Fig. 1. Micrografía mostrando la cercanía de cloroplastos (C), peroxisomas (P) y mitocondrias (M) en una célula vegetal de una planta C3. Micrografía tomada en cooperación con el Dr. Enrique Olmos (CEBAS-CSIC)
¿Qué funciones tiene la Fotorrespiración
El valor adaptativo de la fotorrespiración todavía es materia de debate para muchos fisiólogos vegetales. La fotorrespiración funciona al mismo tiempo que el Ciclo de Calvin-Benson y contribuye a un amplio rango de procesos en el cloroplasto, desde bio-energéticos hasta del metabolismo del C y de asimilación de N. Por ejemplo, el fosfoglicerato se incorpora directamente al Ciclo de Calvin-Benson, mientras que el fosfoglicolato se hidroliza para producir glicolato, que es metabolizado en mitocondrias y en peroxisomas. En el peroxisoma el glicolato se transforma en glicina, que posteriormente es descarboxilada en la mitocondria para producir serina, NH4+ y NADH:

El NH4+ difunde hasta el cloroplasto donde se asimila para formar glutamina (a partir de glutamato), mientras que el NADH puede ser oxidado en la cadena de transporte electrónico de la mitocondria. La serina puede difundir al peroxisoma donde es convertida en glicerato, el cual pasa al cloroplasto para ser fosforilado y formar 3-fosfoglicerato que de nuevo entra en el ciclo de Calvin-Benson.
Como funciones más reconocidas para esta ruta cabe destacar:
1.- Se recupera carbono que podría perderse en forma de 2-fosfoglicerato al principio de la ruta. La fotorrespiración recupera un 75% del carbono perdido como glicerato que de nuevo puede entrar al Ciclo de Calvin-Benson. Es decir, 2 moléculas de 2-fosfoglicerato (4 C) que se pierden en la oxigenación de la Rubisco se transforman en una molécula de 3-fosfoglicerato (3 C). Esta conversión requiere la hidrólisis de una molécula de ATP. Además, la descarboxilación de la glicina en la mitocondria libera una molécula de CO2 que podría llegar al cloroplasto y ser fijada por la Rubisco y entrar en el Ciclo de Calvin.
2.- Otra función atribuida a la fotorrespiración es minimizar la foto-inhibición del aparato fotosintético causado por un exceso de poder reductor formado en el cloroplasto en condiciones de estrés ambiental (alta intensidad luminosa, elevada temperatura, déficit hídrico, salinidad, etc…). Cuando los aceptores electrónicos están saturados, el aparato fotosintético usa sustratos alternativos para eliminar el exceso de poder reductor generado por el flujo electrónico. La fotosíntesis en estas condiciones puede dar lugar a la generación de especies reactivas del oxígeno (ROS) que pueden ocasionar daños oxidativos al cloroplasto y a la célula. La fotorrespiración consume O2 y ATP en el cloroplasto evitando su acumulación. El consumo de O2 en el cloroplasto minimiza su uso como aceptor de electrones en el PSI (reacción de Mehler) o que interaccione con moléculas de clorofila excitadas. Esto minimiza la formación de ROS.
3.- En el estroma del cloroplasto, la glutamina sintetasa y la glutamato sintasa dependiente de ferredoxina (GOGAT) usan el NH4+ y el 2-oxoglutarato, para recuperar el N inicialmente perdido con el glutamato exportado desde el cloroplasto al peroxisoma. En estas reacciones se produce 2 moléculas de Ferredoxina oxidada (Fdox) y dos de ADP, que son empleadas como aceptores electrónicos en la cadena de transporte fotosintético.
En este sentido, la fotorrespiración disipa el exceso de equivalentes reducidos previniendo la sobre-reducción de la cadena de transporte fotosintético.
Cuando se produce un descenso en la relación CO2/O2 intracelular debido al cierre estomático, que se produce en respuesta a estrés hídrico o salino, se suele incrementar el flujo a través de la fotorrespiración.

Fig. 2.- La Fotorrespiración previene la sobre-reducción de la cadena de transporte electrónico en el cloroplasto. La ruta recupera el 75% del C perdido al principio en forma de fosfoglicolato. Además regenera aceptores electrónicos en la cadena de transporte fotosintético (Fdox y ADP) (Imagen tomada de Taiz y Zeiger 2006).
Producción de H2O2 por la Fotorrespiración
En plantas C3 crecidas en condiciones que favorecen una elevada tasa de oxigenación de la Rubisco, tal como un día muy soleado, un estrés hídrico o salino, la ruta fotorrespiratoria es probablemente el proceso más importante y rápido en generar H2O2.
El H2O2 se forma en el peroxisoma durante la oxidación del glicolato a glioxilato por la enzima glicolato oxidasa. En esta situación, la enzima catalasa es crucial para eliminar el H2O2 generado en la fotorrespiración.

La importancia de la catalasa para realizar esta función se ha demostrado con plantas mutantes que presentan baja actividad catalasa o empleando la tecnología antisentido (plantas transformadas con un gen que codifica para la catalasa pero insertado a la inversa provocando así la ausencia del correspondiente mRNA, por lo que no se sintetiza la proteína). Además, se ha demostrado que la catalasa es importantísima para mantener el balance redox celular en condiciones de estrés oxidativo (alta intensidad luminosa, salinidad, ozono etc…) (Willekens et al. 1997).
Aunque los peroxisomas contienen todos los componentes del ciclo ascorbato-glutatión (ASC-GSH) (Jiménez et al., 1997) no es capaz de eliminar la gran cantidad de H2O2 generado en el peroxisoma cuando hay un alto flujo fotorrespiratorio. Sin embargo, una de las funciones atribuidas al ciclo ASC-GSH en el peroxisoma es eliminar el H2O2 que podría difundir por la membrana del peroxisoma y evitar un estrés oxidativo en el citosol (Jiménez et al., 1997, 1998).
La catalasa es un hemoproteína con una alta actividad pero con una baja afinidad (alta Km) por el H2O2. Actúa con altas concentraciones de H2O2, mientras que la APX posee una alta afinidad (baja Km), por lo que es muy activa con bajos niveles de H2O2, actuando en la regulación fina de los niveles intracelulares de H2O2. Esto quiere decir que la eliminación de H2O2 por la catalasa requiere altas concentraciones de esta enzima, mientras que la eliminación por APX requiere menos niveles de enzima. Los altos niveles de enzima catalasa requeridos para esta función se reflejan en los grandes cristales de catalasa que se pueden observar en fotografías de peroxisomas tomadas con microscopio electrónico (Fig 3).

Fig 3. Micrografía mostrando los tres orgánulos implicados en la fotorrespiración, donde se aprecia la localización de la catalasa en la matriz del peroxisoma como una inclusión de forma cuadrada. Imagen sacada del libro “Plant Peroxisomes” (Huang et al, 1983). C, cloroplasto, m , mitocondria, P, peroxisoma).
La generación de H2O2 en la fotorrespiración puede incluso ser mayor que el generado en la reacción de Mehler-peroxidasa (Foyer y Noctor 2003) (ver también https://antioxidantsgroup.wordpress.com/2015/10/06/funcion-del-ascorbato-en-la-proteccion-de-la-fotosintesis-i-la-reaccion-de-mehler-y-el-ciclo-agua-agua/).
Existe una fuerte correlación entre los niveles de actividad catalasa y la tasa fotosintética. La inhibición de la actividad catalasa con el inhibidor aminotriazol hace disminuir la tasa de asimilación de CO2 (Amory et al., 1992). Esto podría ser debido a que el H2O2 es un inhibidor de enzimas que están reguladas por el sistema tiorredoxina como ocurre con algunas enzimas del Ciclo de Calvin-Benson (Gliceraldehido-3-fosfato deshidrogenasa, Fructosa-1,6-bisfosfatasa, Sedoheptulosa-1,7-bisfosfatasa, Fosforibuloquinasa….) (Jacquot et al (1993).
La inhibición de la actividad catalasa puede producirse en condiciones de estrés que afecten a la síntesis de proteínas (salinidad, estrés hídrico, altas y bajas temperaturas etc….) (Vock y Feierabend 1989). Esto tendría un efecto inmediato de acumulación de H2O2 en el peroxisoma, que podría difundir a través de la membrana de este orgánulo y afectar a otros compartimentos celulares.
Conclusiones
Como hemos comentado anteriormente, las funciones adscritas a la fotorrespiración sigue siendo un tema de debate. Cuando se describió este conjunto de reacciones no se le pudo asignar ninguna función útil a la fotorrespiración por diferentes motivos (ver Figura 4):
1.- Se pierde ribulosa-1,5-bisfosfato para el ciclo de Calvin-Benson
2.-La fijación de CO2 se invierte: se consume O2 y se libera CO2 en presencia de luz (de ahí el nombre de fotorrespiración).
3.-Sólo una parte del carbono se recicla
4.- Gasto de ATP de forma innecesaria.

Figura 4.- Flujo del carbono y del oxígeno en el ciclo de Calvin y en la fotorrespiración. (Imagen tomada de Taiz y Zeiger 2006).
Se consideró que la fotorrespiración era una ruta metabólica residual, ineficaz e incluso inútil. Era como si la fotorrespiración deshiciera lo realizado por la fotosíntesis. Pero investigaciones posteriores pusieron de manifiesto que tanto la fotorrespiración como la fotoinhibición (interrupción controlada del PS II) son muy importantes para la regulación de la fotosíntesis. De hecho, mutantes incapaces de hacer fotorrespiración no son capaces de crecer en condiciones ambientales normales, o la supresión del proceso por manipulación genética produce desequilibrios en la planta.
Por tanto, con el paso del tiempo diferentes investigaciones han ido atribuyendo a la fotorrespiración un efecto protector para el proceso de fotosíntesis. Por ejemplo, en situaciones donde la concentración del CO2 es baja. Esta situación ocurre cuando las plantas están creciendo en condiciones de alta intensidad luminosa, cuando hay limitaciones hídricas, salinidad etc… En tales situaciones, un descenso en el proceso de asimilación de CO2 no permitiría una regeneración de los aceptores de electrones en la cadena de transporte fotosintético, especialmente NADP+ y ADP, lo que favorecería la generación de ROS. En estas condiciones, la fotorrespiración es un sumidero alternativo de energía o de poder reductor (Arellano y De la Rivas 2006). Consume, como hemos visto, O2 y ATP en el cloroplasto evitando su acumulación. El consumo de O2 en el cloroplasto minimiza su uso como aceptor de electrones en el PS I (reacción de Mehler; ver también https://cienciacebas.wordpress.com/2015/10/06/funcion-del-ascorbato-en-la-proteccion-de-la-fotosintesis-i-la-reaccion-de-mehler-y-el-ciclo-agua-agua/) o que interaccione con moléculas de clorofila excitadas. Esto minimiza la formación de ROS. Además, permite un flujo electrónico que favorece la recuperación de C para el Ciclo de Calvin-Benson y de aceptores electrónicos (Fdox y ADP).
Es cierto que la fotorrespiración reduce la eficiencia fotosintética (Arellano y De la Rivas 2006), pero ofrece a cambio un mecanismo de fotoprotección en condiciones adversas que opera junto a otros mecanismos de protección, como el ciclo agua-agua y el ciclo de las xantofilas (ver también https://antioxidantsgroup.wordpress.com/2015/10/06/funcion-del-ascorbato-en-la-proteccion-de-la-fotosintesis-i-la-reaccion-de-mehler-y-el-ciclo-agua-agua/ y https://antioxidantsgroup.wordpress.com/2015/10/13/funcion-del-ascorbato-en-la-proteccion-de-la-fotosintesis-ii-los-carotenoides-y-el-ciclo-de-las-xantofilas/).
En consequencia, la Rubisco, enzima sumidero de CO2 o de O2 garantiza un consumo de ATP y NADPH bien mediante el ciclo fotorreductor del carbono (ciclo de Calvin-Benson) o mediante el fotooxidativo (fotorrespiración) (Arellano y De las Rivas 2006).
Bibliografía
Arellano y De las Rivas (2006) Investigación y Ciencia Marzo, pp. 42-50.
Amory et al (1992) Plant Cell Environm 15: 655-663.
Foyer y Noctor (2003) Physiol Plant 119: 355-364.
Huang AHC, Trelease RN, Moore TS Jr (1983) Plant Peroxisomes. Academic Press Inc., New York
Jacquot et al (1993) New Phytol 136: 543-570.
Jiménez et al (1997) Plant Physiol 114: 275-284
Miziorko y Lorimer (1983) Ann. Rev. Biochem. 52:507-535.
Ogren (1984) Ann. Rev. Plant Physiol. 35:415-422.
Taiz y Zieger (2006) Fisiología Vegetal, tercera Edición. Servicio de Publicaciones de la Universidad Jaime I, Castellón (España).
Vock y Feierabend (1989) Plant Cell Environm 12:701-712.
Willekens et al (1997) EMBO J. 16 : 4806-4816.

José A. Hernández es Investigador Científico del Grupo de Biotecnología de Frutales (CEBAS-CSIC)