
Regulación del Cierre Estomático: una Función Representada por Varios Actores
José A. Hernández Cortés (Investigador Científico del CSIC, Grupo de Biotecnología de Frutales, CEBAS)
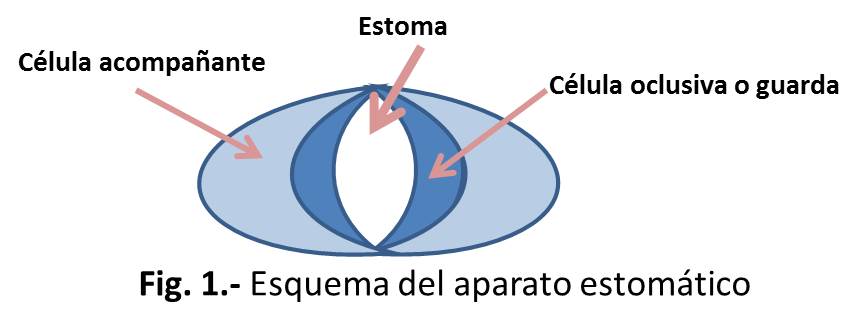
Un estoma es básicamente un poro que es usado para el intercambio gaseoso con el medio y la transpiración de la planta. Está rodeado de dos células, denominadas células oclusivas o guarda, que tienen forma de riñón o de forma de pesa, como ocurre en plantas monocotiledóneas (gramíneas, ciperáceas). Los movimientos estomáticos dependen de cambios en la presión de turgencia de las células oclusivas y las células epidérmicas adyacentes. Al conjunto de células oclusivas y células acompañantes se le denomina aparato estomático (Fig 1).
Las células guarda regulan la abertura de los estomas mediante la integración de diferentes señales, tanto endógenas como exógenas (ambientales). Una adecuada regulación de los estomas va a conseguir un eficiente uso del agua y una tasa óptima de intercambio de CO2 para la fotosíntesis, siendo esencial para una adaptación de las plantas a la falta de agua o estrés hídrico. La habilidad de las plantas para controlar la abertura estomática les permite responder de una forma rápida a los cambios en el medio ambiente. Por ejemplo, puede prevenir una pérdida excesiva de agua, limitar la absorción de compuestos líquidos no deseados o de contaminantes atmosféricos a través de los estomas. Incluso, aún en presencia de abundante agua, las plantas ponen en marcha una regulación temporal de los estomas para limitar las pérdidas de agua durante el proceso de absorción de CO2, es decir, los estomas se abren durante el día y se cierran durante la noche para prevenir pérdidas innecesarias de agua, ya que durante este periodo no se produce fotosíntesis y no hay demanda de CO2. Con los primeros rayos de sol, la planta de nuevo comienza a hacer fotosíntesis, de nuevo los estomas se abren ya que la demanda de CO2 es elevada. La abertura de estomas favorece la transpiración de la planta. Este proceso de transpiración cumple varias funciones:
1.- Se crea una corriente transpiratoria que permite el transporte de nutrientes desde las raíces hasta las partes de la planta en crecimiento.
2.- Enfría las hojas cuando la temperatura es elevada o hay una fuerte intensidad luminosa, es decir, los estomas favorecen una regulación térmica (Sánchez-Díaz y Aguirreolea 2000).
No obstante, algunos investigadores sostienen que, en su conjunto, la transpiración (pérdida de agua) es un mal necesario que entra en conflicto con la necesidad de las plantas para tomar el CO2 necesario para realizar la fotosíntesis.
En condiciones de estrés hídrico, la cadena de transporte electrónico sigue funcionando, lo que favorece que la energía de excitación pueda pasar desde las moléculas de clorofila foto-excitadas directamente al oxígeno, formando oxígeno singlete (1O2). Además, en el fotosistema II se produce la formación de radicales superóxido (O2.-), peróxido de hidrógeno (H2O2) y radicales hidroxilo (.OH)(Suzuki et al 2012). Las células guarda contienen menos cloroplastos y de menor tamaño y un contenido menor de clorofila (entre 1-4%) que las células del mesófilo. Sin embargo, se ha demostrado que el proceso de fotofosforilación (síntesis de ATP en la membrana de los tilacoides) por parte de los cloroplastos de las células guarda es muy activa. Por lo tanto, los ROS generados en la cadena de transporte fotosintética de los cloroplastos de las células guarda puede tener una función muy importante en la respuesta de las plantas a diferentes estímulos ambientales (Pfannschmidt 2003).
Sin embargo, el ácido abscísico (ABA) tiene un protagonismo clave en el control del cierre estomático. El ABA induce la generación de H2O2 en las células guarda dando lugar a un cierre de los estomas (Fig. 2). La generación de H2O2 inducida por ABA tiene dos posibles fuentes: (1) los cloroplastos y (2) la actividad NADPH oxidasa de membrana plasmática (Zhang et al 2001a; 2001b). La NADPH oxidasa de membrana es una enzima que reduce el oxígeno a O2.-, que posteriormente será dismutado hasta H2O2 por acción de la actividad SOD. En Arabidopsis, la NADPH oxidasa presenta dos subunidades (AtrbohD y AtrbohF). En este sentido, un doble mutante atrbohD/F mostró un reducido cierre estomático así como una reducida producción de especies reactivas de oxígeno (ROS) comparado con las plantas silvestres. La aplicación exógena de H2O2 restauraba el cierre estomático, lo que revelaba una conexión entre la generación de ROS por la NADPH oxidasa y el ABA en el cierre estomático (Kwak et al. 2003).

Está bien documentado que la concentración de iones determina la turgencia de las células guarda y el tamaño de la abertura estomática. Los cambios de turgencia pueden ser debidos a una modificación del potencial hídrico (ψ) de las células oclusivas o de cambios activos del potencial osmótico (ψs). Ambos mecanismos conllevan movimientos de agua en ambas direcciones en las células guarda, que finalmente va a producir una abertura o cierre del estoma. Cuando el estoma se abre se produce un aumento masivo de solutos en las células guarda. Esto provoca un descenso de ψs y una entradas de iones, sobre todo K+ (también Na+ en plantas halófitas) y Cl– hacia las células guarda u oclusivas. La entrada de K+ se produce por el bombeo activo de H+ fuera de las células oclusivas por acción de una H+-ATPasa de membrana. La entrada de K+ se equilibra con una entrada de Cl– y sobre todo con la acumulación del ion malato. El ácido málico se produce a partir del ácido oxalacético, que a su vez procede del catabolismo del almidón almacenado (Sánchez-Díaz y Aguirreolea 2000). El estoma se cierra cuando la H+-ATPasa se inactiva y el K+ y el Cl– salen de forma pasiva de las células oclusivas. Por otro lado, el aumento de los niveles intracelulares CO2 favorece la salida de algunos aniones (Cl- y malato) de las células guarda, lo que también contribuye al cierre de los estomas (Fig. 2).
Se ha demostrado que el H2O2 inactiva el transporte de K+ hacia el interior celular y produce una alcalinización del citosol al activar el transporte de H+ hacia la vacuola. Al mismo tiempo, el H2O2 puede mediar en la señalización del ABA al activar los canales de Ca2+, aumentando su concentración en el citosol y activando el cierre de los estomas (Pei et al. 2000). Un estudio reciente ha demostrado que el receptor del ABA (PYR/PYL/RCAR) regula los canales de K+ y de Cl– mediante la activación de los canales de Ca2+ de membrana plasmática por parte de las ROS (Wang et al 2013).
Las ROS pueden actuar sobre las MAPKs (protein kinasas activadas por mitógeno), MAPK fosfatasas o proteín kinasas dependientes de Ca2+ modulando su actividad. EL H2O2 inactiva las protein fosfatasas ABI1 y ABI2 (Meinhard et al 2002), como también ocurre para la ABI1 con la unión del ABA a su receptor, liberándose una proteína (OST1) que actúa fosforilando y activando la NADPH oxidasa de membrana, incrementando la producción de H2O2 (Kepka et al., 2011). EL H2O2 también actúa como un segundo mensajero, de modo que puede activar algunas MAPKs que actúan promoviendo el cierre estomático (Jiang et al., 2008).
Como hemos visto, el cierre de los estomas no depende de una sola señal, sino que se establece una red de comunicación por parte de diferentes actores, siendo los más importantes el ABA, la NADPH oxidasa de membrana, el H2O2, y otros como las MAPKs, transportadores de aniones y cationes etc… (Fig. 2), que responde de una forma coordinada a estímulos externos y que finalmente van a favorecer que la planta se adapte de la mejor forma posible a situaciones diversas de estrés.
Bibliografía empleada: Jiang et al. 2008, Plant Cell Reports 27: 377-385. Kepka et al. 2011, Plant Physiol 157:2108-2119 Kwak et al. 2003, EMBO J 22: 2623-2633. Meinhard et al. 2002, Planta 214:775-782. Pei et al. 2000, Nature 406: 731-734. Pfannschmidt 2003, Trends Plant Sci 8: 33-41. Sánchez-Díaz y Aguirreolea 2001, Fundamentos de Fisiología Vegetal (Azcón J., Talón M., Eds.) Suzuki et al. 2012, Plant Cell Environm 35: 259-270. Wang et al. 2013, Plant Physiol 163: 566-577 Zhang et al. 2001a, Cell Reseach 11:195-202; Zhang et al. 2001b, Plant Physiol 126: 1438-1448.
